
基本介紹
- 中文名:厭氧氨氧化菌
- 外文名:Anaerobic ammonia oxidant
- 個體形態特徵:呈球形、卵形
- 菌屬:革蘭氏陰性菌
- 莢膜:無
- 重要意義:全球氮循環;污水處理
基本介紹,特性,個體形態特徵,化學組分特徵,菌屬分類,生化反應機理,可能反應機理,厭氧氨氧化,套用,氧化工藝,聯合工藝,工藝,耦合,
基本介紹
厭氧氨氧化(anaerobic ammonium oxidation, Anammox)菌為自養型細菌,可在缺氧條件下以氨為電子供體,亞硝酸鹽為電子受體,產生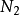 。已發現的厭氧氨氧化菌均屬於浮霉狀菌目(Planctomycetales)的厭氧氨氧化菌科(Anammoxaceae),共 6 個屬,分別為 Candidatus Brocadia、Candidatus Kuenenia、Candidatus Anammoxoglobus、CandidatusJettenia、Candidatus Anammoximicrobium moscowii 及 Candidatus Scalindua。其中,Candidatus Scalindua 發現於海洋次氧化層區域,稱之為海洋厭氧氨氧化菌,其餘 5 個屬均發現於污水處理系統中,稱之為淡水厭氧氨氧化菌。厭氧氨氧化細菌對全球氮循環具有重要意義,也是污水處理中重要的細菌。
。已發現的厭氧氨氧化菌均屬於浮霉狀菌目(Planctomycetales)的厭氧氨氧化菌科(Anammoxaceae),共 6 個屬,分別為 Candidatus Brocadia、Candidatus Kuenenia、Candidatus Anammoxoglobus、CandidatusJettenia、Candidatus Anammoximicrobium moscowii 及 Candidatus Scalindua。其中,Candidatus Scalindua 發現於海洋次氧化層區域,稱之為海洋厭氧氨氧化菌,其餘 5 個屬均發現於污水處理系統中,稱之為淡水厭氧氨氧化菌。厭氧氨氧化細菌對全球氮循環具有重要意義,也是污水處理中重要的細菌。
特性
在厭氧氨氧化過程中,羥胺和肼作為代謝過程的中間體。和其它浮黴菌門細菌一樣,厭氧氨氧化菌也具有細胞內膜結構,其中進行氨厭氧氧化的囊稱作厭氧氨氧化體(anammoxosome),小分子且有毒的肼在此內生成。厭氧氨氧化體的膜脂具有特殊的梯烷(ladderane)結構,可阻止肼外泄,從而充分利用化學能,且避免毒害。
個體形態特徵
厭氧氨氧化菌形態多樣,呈球形、卵形等,直徑0.8-1.1μm。厭氧氨氧化菌是革蘭氏陰性菌。細胞外無莢膜。細胞壁表面有火山口狀結構,少數有菌毛。.細胞內分隔成3部分:厭氧氨氧化體(anammoxosome)、核糖細胞質
(riboplasm )及外室細胞質(paryphoplasm ) 。核糖細胞質中含有核糖體和擬核,大部分DNA存在於此。厭氧氨氧化體是厭氧氨氧化菌所特有的結構,占細胞體積的50%-80%,厭氧氨氧化反應在其內進行。厭氧氨氧化體由雙層膜包圍,該膜深深陷入厭氧氨氧化體內部。
化學組分特徵
厭氧氨氧化菌的細胞壁主要由蛋白質組成,不含肽聚糖。細胞膜中含有特殊的階梯烷膜脂,由多個環丁烷組合而成,形狀類似階梯。在各種厭氧氨氧化菌中,階梯烷膜脂的含量基本相似。疏水的階梯烷膜脂與親水的膽鹼磷酸、乙醇胺磷酸或甘油磷酸結合形成磷脂,構成細胞膜的骨架。細胞膜中的非階梯烷膜脂由直鏈脂肪酸、支鏈脂肪酸、單飽和脂肪酸和三萜系化合物組成。曾一度認為階梯烷膜脂只存在於厭氧氨氧化體的雙層膜上,其功能是限制有毒中間產物的擴散。目前認為階梯烷膜脂存在於厭氧氨氧化菌的所有膜結構上(包括細胞質膜) ,它們與非階梯烷膜脂相結合,以確保其他膜結構的穿透性好於厭氧氨氧化體膜。
菌屬分類
(1)Candidatus“Brocadia anammoxidans”
Strous等研究了SBR反應器中厭氧氨氧化污泥(優勢菌為B.anammoxidans)的生理學參數。富集培養得到的細菌70%以上是一種優勢自養菌。經過pH7.4、20 mmol/L的K2HPO/KH2P04緩衝劑和2.5%的戊二醛混合液處理後,在電子顯微鏡下表現出不規則的微生物特性。採用改進的Percoll密度梯度離心分離法分離得到了非常純的細胞懸浮物,在每200~800個細菌中含有不到1個雜細菌。這些純化的厭氧氨氧化菌活性很高,從中提取的DNA通過PCR擴增和1 6S rRNA的分析,證明B.anammoxidans是厭氧氨氧化的功能微生物,確認了厭氧氨氧化菌是Planctomycetales序列中自養菌的一個新成員,被命名為CandidatuJ“B.anammoxida”。
(2)Candidatus “Kuenenia stuttgartiensis”
Candidatus“K.stuttgartiensis”菌是從德國和瑞士幾個污水處理廠的生物膜反應器中發現的。Egli等的研究結果表明,k.stuttgartiensis 與B.anammoxidans的作用方式類似,電鏡顯示其細菌結構也類似。但它對磷酸鹽有更高耐受性(20 mmol/L),對亞硝酸鹽耐受性則為
13 mmol/L,在細胞密度更低的情況下有活性。最大的厭氧氨氧化活性(以單位蛋白質計)為26.5 nmol/(mg·min),比B.anammoxidans低。pH範圍是6.5~9.0,最佳為8.0,最佳溫度為37℃。研究發現,當溫度升至45℃時觀察不到厭氧氨氧化活性,並且當溫度回降至37℃時厭氧氨氧化活性不恢復;在ll℃時的活性約為37℃的24%,可見它適宜生存於中溫偏鹼性環境。
(3)Candidatus“Scalindu sorokinii”
最近在黑海發現的Candidatus“Scalindua sorokinii”。它又分為兩種厭氧氨氧化菌種——Candidatus “Scalindua brodae”和Candidatus “Scalindua wagneri”,其生理特性研究較少。(4)Candidatus“Anammoxoglobus propionicus”
Candidatus“Anammoxoglobus propionicus”能代謝丙酸鹽其生理特性研究較少。
生化反應機理
可能反應機理
Van de Graaf等用N作為示蹤元素,研究了厭氧氨氧化代謝途徑。他們根據N2H4轉化為N2的過程給N02還原為NH20H的反應提供等量電子的假設。提出了兩種可能的機理。其一,一個由膜包圍的酶複合體將氨和NH2OH轉化為N2H4,N2H4則在外周胞質內氧化為氮氣,產生的電子通過內部電子轉移,在包含酶複合體(此酶複合體也負責N2H4氧化)的細胞質中將N02還原為NH2OH。其二,氨和NH2OH在細胞質內被一由膜包圍的酶複合體轉化為N2H4,
N2H4在外周胞質內轉化為N2,與產生的電子通過電子傳輸鏈傳遞給細胞質內的亞硝酸鹽還原酶將N02還原為NH2OH。
厭氧氨氧化
根據厭氧氨氧化反應的關鍵酶是位於厭氧氨氧化體中的肼氧化酶(HZO)的觀點,提出了與厭氧氨氧化體膜相關的生化模型,NH4和羥胺(NH2OH)被肼水解酶(HH)轉化為肼,肼又被肼氧化酶(HZO)氧化,HZO與HAO(N.europaea)相似。肼的氧化發生在厭氧氨氧化體的內部,形成N2、4個質子和4個電子。這4個電子與來自核糖質中的5個質子一起通過亞硝酸還原酶(NIR)將亞硝酸鹽還原為羥胺。在這個模型中,通過在核糖質中的質子消耗和在厭氧氨氧化體裡面的質子產生,厭氧氨氧化反應建立了一個質子梯度。這就在厭氧氨氧化體和核糖質之間產生了電化學質子梯度。這種梯度包含有化學勢能(△pH)和電子勢能。
化學勢能和電子勢能產生使質子從厭氧氨氧化體裡面移動到厭氧氨氧化體外面的一種質子驅動力△p。在厭氧氨氧化體膜束縛三磷酸腺苷酶(ATPase)的催化作用下合成三磷酸腺苷(ATP)。質子通過三磷酸腺苷酶形成的質子孔被動遷移回到核糖質中,厭氧氨氧化體膜束縛
三磷酸腺苷酶位於核糖質中球形親水的ATP合成區和厭氧氨氧化體膜中非親水的質子遷移區,合成的ATP釋放在核糖質中。
套用
氧化工藝
Mulder等在厭氧流化床中發現了厭氧氨氧化。後來,Van de Graaf等和Bock等發現了以亞硝酸鹽為電子受體的厭氧氨氧化過程。鄭平等研究了厭氧氨氧化菌混培物的動力學特性[141。Fux Christian等進行中試試驗研究,首先在連續攪拌反應器中完成氨氧化,58%的NH4-N轉化為NO2;在SBR中完成厭氧氨氧化,除N速率為2.4 kg/(m·d),除N率達90%;Sliekers等在氣提式反應器中發現除N速率達8.9 kg/(m·d),這個除N速率是實驗室所獲得的除N速率的20倍。Dapena-Mora等研究中發現在氣提式反應器中N負荷率為2.0 g/(L·d),最大比厭氧氨氧化活性(MSAA)為0.9 g/(g·d);在SBR中N負荷率為0.75 g/(L·d),MSAA為0.4 g/(g·d),除N02率達99%。
聯合工藝
Jetten等利用SHARON-ANAMMOX聯合工藝對污泥消化出水進行了研究。SHARON反應器總氮負荷為0.8 kg/(m·d),轉化53%的總氮(39%NO2,14%N03),用SHARON反應器的出水作為厭氧氨氧化流化床反應器的進水,在限制N02的厭氧氨氧化反應器中N02全部被除去,試驗中NH4-N的去除率達83%。Van Dongen等套用SHARON-ANAMMOX聯合工藝在工廠中長時間穩定運行。
工藝
Dijkman和Strous描述了一個新的生物脫氮工藝CANON,在限氧條件下(<0.5%空氣飽和度)得到了好氧和厭氧氨氧化茵的混培物,NH4被需氧氨氧化菌(Nitrosomonas和Nitrososira)氧化為亞硝酸鹽,然後被厭氧氨氧化菌轉化為氮氣,此過程依賴於2種白養微生物菌群(Nitrosomonas需氧菌和Planctomycete厭氧氨氧化菌)的協同作用。CANON在2種不同的反應器(SBR和恆化器)中進行了研究,容積負荷(N)0.1 kg/(m·d),除氮達92%。Sliekers等發現在限氧條件以及好氧氨氧化菌和厭氧氨氧化菌都有合適的負荷率時,SBR反應器中除N負荷率達0.3 kg/(m·d),NH4主要轉化為N2(85%),其餘的轉化為硝酸鹽(15%)。Sliekers等用氣提式反應器,除N負荷率達1.5 kg/(m·d),這個速率是以前實驗室獲得的速率的20倍。Hao等[181開發了在生物膜反應器中混合硝化(氨氧化+亞硝酸鹽氧化)、厭氧氨氧化的數學模型,評價了CANON過程的溫度、流速。
耦合
Jetten等通過污泥消化產甲烷除去COD,N部分氧化至NO2,然後以NH4為電子供體反硝化,實現了甲烷化和厭氧氨氧化。Zhang運用EGSB反應器技術,COD的去除率97%,N02去除率100%,容積負荷達6.56 g/(L·d)(COD)和0.99/(L·d)(N),實現了甲烷化、反硝化與厭氧氨氧化的耦合。
