貯存在地殼裡的礦物元素,通過自然風化、沉積物的分解和人類的開採冶煉轉變為可被生態系統利用的營養物質,為植物所吸收,參與生命物質的形成,並沿食物鏈轉移。然後,由動植物殘體或排泄物經微生物的分解作用,將元素返迴環境的過程。
基本介紹
- 中文名:沉積型循環
- 外文名:sedimentary cycle
- 性質:蓄庫主要是岩石圈和土壤圈
- 營養元素:磷、硫、鉀、鈉、鈣等
簡介,硫的循環,磷的循環,
簡介
沉積型循環(sedimentary cycle)的蓄庫主要是岩石圈和土壤圈。屬於沉積型循環的營養元素主要有磷、硫、鉀、鈉、鈣等。保存在岩石圈中的這些元素只有當地殼抬升變為陸地後,才有可能因岩石風化、侵蝕和人工採礦等形式釋放出來被生產者植物所利用。因此,循環周期很長,常常還會造成局部性的匱乏。
硫的循環
硫雖然也有氣態化合物(如二氧化硫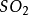 ),但它對硫的循環所起的作用很小,在硫的循環過程中,比氣體型循環有更多的停滯階段,其中海洋和大陸深水湖的沉積層就是最明顯的停滯階段。雖然少數生物可以從胺基酸(有機硫)中獲得它們所需要的硫,但大多數生物都是從無機的硫酸鹽(SO42-)中獲得它們所需要的硫。通過生物學過程所合成的硫氫基(-SH),在分解過程中大部分都能被麴黴屬(Aspergillus)和脈孢菌屬(Neurospora)中的真菌和細菌所礦化。在厭氧條件下還可直接被屬於埃希氏桿菌屬(Escherichia)和變形桿菌屬(Proteus)的細菌降解為硫化物,也可被降解為元素硫或硫化物,能還原硫酸鹽的厭氧菌都是異養細菌。
),但它對硫的循環所起的作用很小,在硫的循環過程中,比氣體型循環有更多的停滯階段,其中海洋和大陸深水湖的沉積層就是最明顯的停滯階段。雖然少數生物可以從胺基酸(有機硫)中獲得它們所需要的硫,但大多數生物都是從無機的硫酸鹽(SO42-)中獲得它們所需要的硫。通過生物學過程所合成的硫氫基(-SH),在分解過程中大部分都能被麴黴屬(Aspergillus)和脈孢菌屬(Neurospora)中的真菌和細菌所礦化。在厭氧條件下還可直接被屬於埃希氏桿菌屬(Escherichia)和變形桿菌屬(Proteus)的細菌降解為硫化物,也可被降解為元素硫或硫化物,能還原硫酸鹽的厭氧菌都是異養細菌。
在厭氧環境中(主要是在水生生態系統的深層)硫化氫的大量存在對大多數生物來說都是有害的,200米以下的黑海海域缺乏較高等的動物可能與此有關。
化石燃料的不完全燃燒可使二氧化硫()進入大氣圈,這是大氣遭受污染的一個主要原因。大氣中的氧化硫、二氧化硫和元素硫可被進一步氧化形成三氧化硫( ),它與水結合便形成了硫酸(
),它與水結合便形成了硫酸( ),雨水中含有硫酸就會形成酸雨。
),雨水中含有硫酸就會形成酸雨。
自然界的生態平衡機制可使一種生物的毒性被另一種生物所處理。無色的硫細菌[如貝氏硫細菌(Beggiatoa)]可把硫化氫氧化為元素硫,而硫桿菌(Thiobacillus)則可把它氧化為硫酸鹽。有些化能合成細菌可利用氧化過程中釋放出的能量來還原二氧化碳並從中獲得它們所需要的碳:

這些細菌也包括綠色和紫色光合作用細菌,它們在還原二氧化碳的過程中可利用硫化氫中的氫作為氧的受體。綠色細菌只能使硫化物氧化為元素硫,而紫色細菌則能使氧化進行到硫酸鹽的階段:
硫循環的沉積過程往往與鐵離子和鈣離子的存在有關,當形成不溶性的FeS、 和硫酸鈣
和硫酸鈣 時就會出現硫的沉澱現象。其中的FeS具有重要的生態學意義,它是在厭氧條件下生成的,在中性和鹼性水中呈不溶解狀態。因此,在這種條件下硫的存在就成了鐵含量的限制因素。由於FeS的熱力學過程,其他一些在生物系統中很重要的營養物質如銅、鎘、鋅和鈷等也可能在一定時期內受到限制。另一方面,正是由於這些鐵化合物的粘合作用,也可使磷從不溶解形態轉化為可溶解形態,因而增加了磷的可利用性。在厭氧的含硫沉積底泥中,硫酸鹽和硫化物的還原過程在很大程度上控制著整個生態系統的生物化學過程:這說明在不同的礦物質循環之間存在著相互作用和調節現象,而在這些循環的內部也存在著複雜的生物學和化學調節。
時就會出現硫的沉澱現象。其中的FeS具有重要的生態學意義,它是在厭氧條件下生成的,在中性和鹼性水中呈不溶解狀態。因此,在這種條件下硫的存在就成了鐵含量的限制因素。由於FeS的熱力學過程,其他一些在生物系統中很重要的營養物質如銅、鎘、鋅和鈷等也可能在一定時期內受到限制。另一方面,正是由於這些鐵化合物的粘合作用,也可使磷從不溶解形態轉化為可溶解形態,因而增加了磷的可利用性。在厭氧的含硫沉積底泥中,硫酸鹽和硫化物的還原過程在很大程度上控制著整個生態系統的生物化學過程:這說明在不同的礦物質循環之間存在著相互作用和調節現象,而在這些循環的內部也存在著複雜的生物學和化學調節。
磷的循環
磷是比較典型的沉積型循環物質,這種類型的循環物質實際上都有兩種存在相:岩石相和溶鹽相。這類物質的循環都是起自岩石的風化,終於水中的沉積。岩石風化後,溶解在水中的鹽便隨著水流經土壤進入溪、河、湖、海並沉積在海底,其中一些長期留在海里,另一些可形成新的地殼,風化後又再次進入循環圈。動植物從溶鹽中或其他生物中獲得這些物質,死後又通過分解和腐敗過程而使這些物質重新回到水中和土壤中(圖5-40)。
磷的主要儲存庫是天然磷礦。由於風化、侵蝕作用和人類的開採活動,磷才被釋放出來。一些磷經由植物、植食動物和肉食動物而在生物之間流動,待生物死亡和分解後又使其重返環境。在陸地生態系統中,磷的有機化合物被細菌分解為磷酸鹽,其中一些又被植物吸收,另一些則轉化為不能被植物利用的化合物。陸地的一部分磷則隨水流進入湖泊和海洋。
在淡水和海洋生態系統中,磷酸鹽能夠迅速地被浮游植物吸收,而後又轉移到浮遊動物和其他動物體內。浮遊動物每天排出的磷量約與其生物量中所儲存的磷量相等,從而使循環持續進行。浮遊動物排出的磷有一半以上是可以被浮游植物吸收的無機磷酸鹽。水體中其他的有機磷可被細菌利用,細菌又被一些小動物取食,這些小動物可以排泄磷酸鹽。一部分磷沉積在淺海,一部分磷沉積在深海。一些沉積在深海的磷又可以隨著海水的上涌被帶到光合作用帶並被浮游植物利用。由於動植物殘體的下沉,常使水錶層的磷被耗盡而深水中的磷過多。
人類的活動已經改變了磷的循環過程。由於農作物耗盡了土壤中的天然磷,人們便不得不施用磷肥。磷肥主要來自磷礦、魚粉和鳥糞。由於土壤中含有許多鈣、鐵和銨離子,大部分用作肥料的磷酸鹽都變成了不溶性的鹽而被固結在土壤中或池塘、湖泊及海洋的沉積物中。由於很多施於土壤中的磷酸鹽最終都被固結在深層沉積物中,並且由於浮游植物不足以維持磷的循環,所以沉積到海洋深處的磷比增加到陸地和淡水生態系統中的磷還要多。
用放射性同位素32P標誌海洋浮遊動物的試驗表明:磷酸鹽的排泄速率與動物的呼吸率成正比,這就是說,磷的周轉時間是直接與代謝率相關的。由於代謝率是動物體積的負函式,因此物質的周轉率便隨動物體積的增大而降低。作為分解者的微生物具有很高的物質周轉率,而作為頂位肉食動物的大動物則只有較低的物質周轉率。Whittaker曾研究過放射性磷在池塘中的移動,他發現生物越大32P的吸收越少,磷的周轉率越慢。較大的生物對於放射性磷的積累一般需要較長的時間,而且磷的累積濃度也低於體積小的生物。大生物體內磷的消失速率也比較慢,由於大生物體內具有穩定的物質利用系統,因此對來自環境物質的影響在一定程度上具有緩衝能力。
在很多情況下,物種在物質循環中所發揮的作用遠遠超過它們在能流方面對生態系統的貢獻。例如,棲息在河口潮間帶的肋貽貝,每天從海水懸浮顆粒中移走大約三分之一的磷,更確切地說,每2.6天便能使海水懸浮顆粒中的磷周轉一次,這些磷的大部分又被肋貽貝吐出,沉澱在底泥上而被以底泥為食的動物所利用,這些動物可以把磷酸鹽再釋放到生態系統中去。肋貽貝在磷的循環中起著重要的作用,然而它在生態系統能流中的作用則是微不足道的。可見一種生物在生態系統中的重要性不能總是以其在能流中的功能來衡量的。
